体内受精環境に対応する精子―卵相互作用の適応進化に関する研究
体内受精は、陸上環境に適応したほ乳類、鳥類、は虫類などの脊椎動物や昆虫類に共通した受精様式です。両生類は、脊椎度物の進化において水中から陸上に進出した最初の動物群ですが、現存種に見られる生殖様式は水中や樹上での体外受精や体内受精など、極めて多様です(Duellman & Trueb ,1986)。このことは、水中や陸上の様々な受精環境への受精様式の適応と体内受精の進化が両生類の陸上進出に大きく貢献したことを意味します。両生類は種間交雑が多くの種間で可能であり、受精システムは基本的に共有されていると考えられます。一方で、これまでに発見された5000を越える種の中で見られる様々な生殖様式の存在は、両生類の受精に未知のメカニズムが機能している可能性を示しています。例えば、ツメガエルなど、多くの無尾両生類は淡水中で放卵・放精し、精子は淡水に触れると運動を開始して卵に到達し、受精します。淡水による精子運動の開始は淡水魚にも見られるメカニズムです。しかし、無尾両生類と共通の祖先種から分岐した有尾両生類ではほとんどの種が体内受精を行い、精子と卵は受精時に淡水に曝されることはありません。実際、日本を代表する有尾両生類であるアカハライモリの体内受精では、メスの輸卵管の出口付近の貯精嚢に蓄えられた精子は、輸卵管を通過してきた卵に媒精され、卵は受精した後水中に産み落とされます(図1)。体内環境で受精するためには淡水以外の要因で精子運動を開始させるメカニズムが必要です。
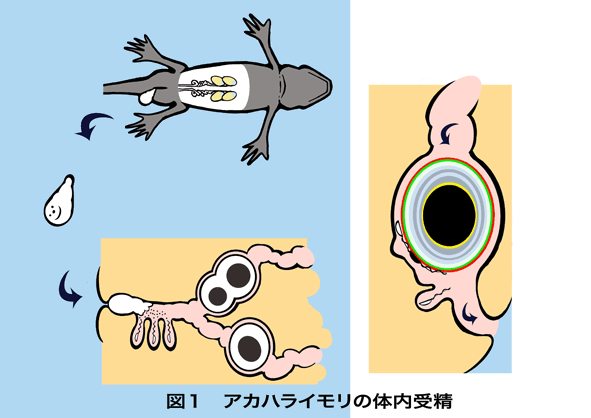
図1 アカハライモリの体内受精
両生類の受精システムは主に淡水中で体外受精をする無尾両生類で研究されてきました。しかし、両生類にはイモリなどの有尾両生類、アシナシイモリなどの無足両生類が存在し、イモリに見られるような体内受精様式やアシナシイモリに見られるような胎生様の受精様式等が発達しています。当研究室ではアカハライモリの体内受精機構について、精子運動調節や先体反応誘起調節、精子と卵膜の結合等、卵-精子相互作用の分子メカニズムを研究しています。アカハライモリに特有の分子メカニズムを通して、体外から体内へ受精環境の変化に対応する受精システムの変化を可能にした精子の分子基盤の解明を目指しています。
(1) 両生類における受精様式の進化
Duellman & Trueb (1986) は、両生類の様々な生殖様式を類型化して、両生類の形態的な進化との関連を示しました(図2は体内受精の進化に関するものを抜き出しています)。有尾両生類では、早い時期に分化したサンショウウオなどは体外受精を行いますが、進化が進んだイモリなど、多くの種では体内受精を行います。一方、
無尾両生類では、ほとんどの種が体外受精を行いますが、ヒキガエル科などの数種で体内受精が報告されています。無足両生類は、進化的に早く分岐したグループですが、このグループに属する種は知られている限りすべてが体内受精を行います。このように、現生両生類では体内受精様式を行う種と体外受精を行う種がそれぞれ大きなグループを形成しています。
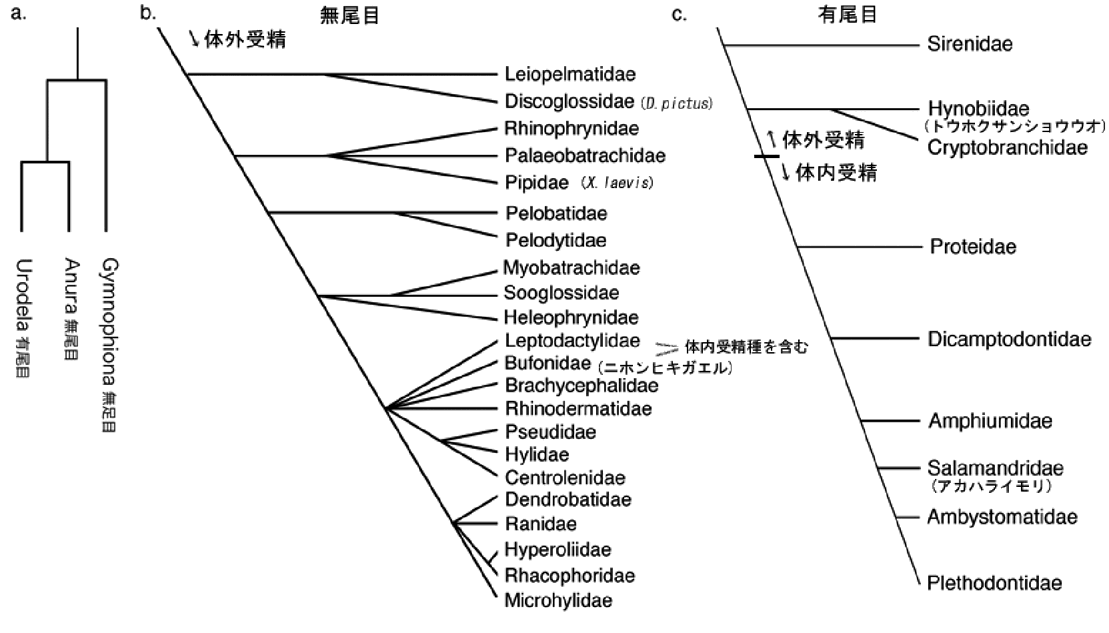
図 2 両生類における体内受精の進化
(2) アカハライモリの体内環境に特異的な受精過程(体外受精を行うツメガエルとの比較)
受精は精子の活性化に始まり精子と卵の合一に至る現象です。受精はその過程で起こる精子選択や精子競争によって、種特異性など、特有の性質を持っています。受精過程の初期に見られる精子-卵相互作用は、卵細胞を取り巻く卵外被と精子との間で起こる現象で、卵と精子を取り巻く外部環境に適応した多様な信号伝達を介して成し遂げられます。図3は、両生類の2種類の卵外被(ジェリー層と卵膜)において、精子-卵相互作用が起こる順序を、アカハライモリとツメガエルについてまとめた模式図です。ツメガエルでは、精子はメスを抱接したオスによって淡水中に放出されると、低浸透圧に反応して運動を開始します(Inoda & Morisawa, 1987)。ジェリー層中の誘因物質によってジェリー層にたどり着いた精子はジェリー層中に侵入し、ジェリー層の構造による排除を受けつつ卵膜に達し、先体反応を起こして卵膜に結合します。一方、アカハライモリでは、精子は体内で受精するため、上述のように淡水に換わる精子運動の引き金があるはずです。また、精子は直接卵に媒精されるため、卵への誘引は必要ではないでしょう。少なくとも、ツメガエルで知られている精子誘引物質はアカハライモリでは合成されていません。さらに、先体反応はジェリー層で起こり (Sasaki et al., 2002)、ジェリー層の構造は精子を排除するのではなく、むしろ卵に導く物理的なガイドとして受精の効率を高めます (Itoh et al., 2002)。精子は卵膜と結合しますが、結合を担うタンパク質はツメガエルとは異なっているようです(Nakai et al., 1999)。アカハライモリの受精に見られるこのような特徴の多くは体内環境下での受精の成立に寄与するものと推察されます。私たちは、特に先体反応と精子運動に焦点を当てて、アカハライモリ精子-卵相互作用の調節機構を調べ、精子運動を開始させるタンパク質であるSperm Motility- Initiating Substance(SMIS)と先体反応を誘起するタンパク質であるAcrosome Reaction-Inducing Substance (ARIS) がジェリー層に存在することをつきとめました。
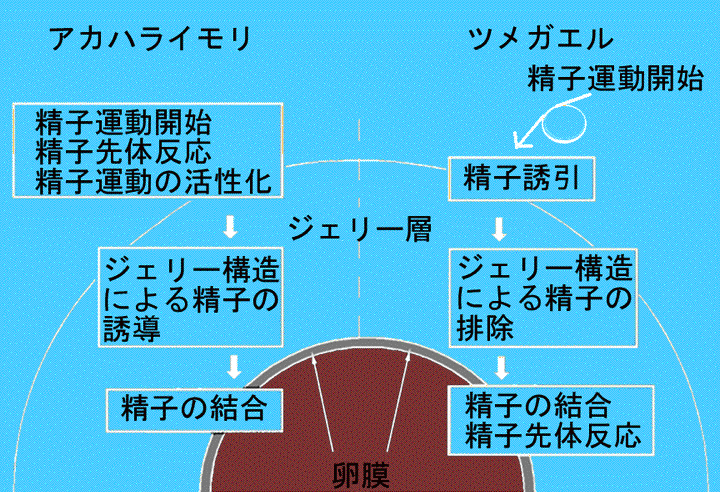
図 3 アカハライモリの卵(左)と精子-卵相互作用の比較(右)
(3)先体反応と連動する精子運動開始機構
先体反応は、イロワケガエル(Campanella et al., 1997)やイベリアトゲイモリ(Picheral,1977)ではジェリー層、ヒキガエル(Yoshizaki & Katagiri,
1982)では卵膜と、両生類の中でも種によって誘起される部位が異なります。上述のようにアカハライモリの精子はジェリー層で先体反応を起こしますが、輸卵管から6層のジェリー層の付加過程にある卵を採取して、精子を媒精し、先体反応と受精率との関係を調べたところ、アカハライモリではジェリー層の表面で先体反応が起こることが受精成立に重要であることが明らかになりました(Takahashi et al., 2006;図4)。
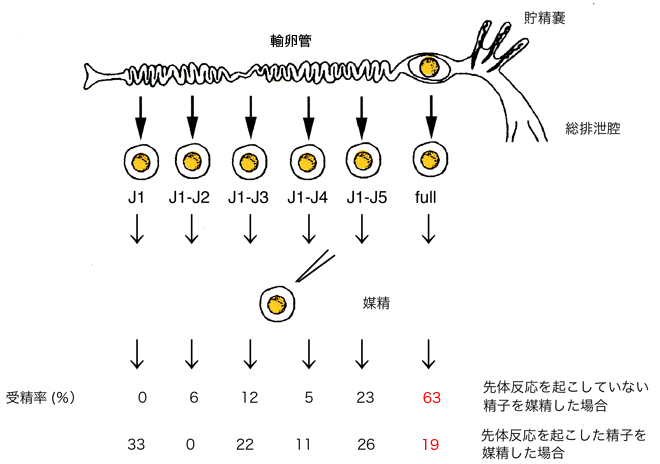
図4 アカハライモリ卵の媒精実験
先体反応をあらかじめ誘起した精子では十分に受精が起こらない。
一方、アカハライモリのARISとSMISに 特異的に結合する抗体を作成し、ARISとSMISの特徴を調べたところ、SMISは分子量が約34kDaの糖鎖を持たないタンパク質であり、ARISはN型及びO型糖鎖を持つ分子量約80 kDa及び122 kDaの糖タンパク質であることが明らかになりました(Watanabe et al., 2009; Watanabe et al., 2010)。また、SMISはジェリー層表面で顆粒状の構造に分布しており、ARISが分布するシート状の構造によって覆われて外部環境から隔離されていました(図5)。従って、メスの体内でジェリー層表面に媒精された精子では、まずARISによって先体反応が誘起され、これによってシート状構造を崩壊させることで初めてSMISの信号を受けることになります (Watanabe et al., 2010; Yokoe et al., 2014)。先体反応がジェリー層表面で起こることの重要性はこのような作用機序と関わるものと考えられます。一般に先体反応は精子運動が開始・活性化された後に起こることから、「先体反応誘起と連携した精子運動の開始機構」は大変ユニークですが、アカハライモリの体内受精には必須の機構です。

図5 ジェリー 層表面でのARISとSMISの局在(模式図)
赤: ARIS、緑: SMIS、黄: 精子先体中の分解酵素。
(4)システインノットタンパク質、SMIS
SMISに特異的な抗体によって検出されたジェリー層中の34kDaのタンパク質を生化学的、及び分子生物学的に解析したところ、SMISは150個のアミノ酸で構成されたシステインノットタンパク質であることがわかりました (Yokoe et al.,
2016)。システインノットは、タンパク質の二量体・多量体形成に関わる機能的な構造の名称です。SMISは精子の中片に結合するアミノ酸配列を、システインノットによって形成される3つのループのうちの第2ループに持っています。また、その他のループはSMISやジェリー層に含まれる他のタンパク質との重合に関り、SMISが局在する顆粒構造の形成に寄与すると考えられます。動物の精子の運動は、水中の浸透圧やpH、卵に由来するアミノ酸やステロイド、ペプチド、タンパク質など、種によって様々な要因によって調節されますが、SMISは初めて発見された精子運動を調節するシステインノットタンパク質です。SMISはこれまで報告されているシステインノットタンパク質との相同性が低い新奇タンパク質で、ジェリー層成分を合成・分泌する輸卵管でだけで合成・分泌されていることから、受精のために新たに獲得された遺伝子にコードされていると考えられます。
(5)SMISによる信号伝達の特徴
ジェリー層中のSMISは精子に作用するために活性化される必要があります (Mizuno et al., 1999; Watanabe et al., 2010)。これは、これまでに報告されている精子運動調節タンパク質と異なり、SMISに特有の特徴です。SMISの活性化機構はよくわかっていませんが、先体反応によって放出される酵素によって活性化されることが推察されています(Yokoe et al., 2014)。活性化されたSMISは精子中片に結合することが示唆されており、SMIS受容体は中片に存在すると考えられます。アカハライモリの精子では、細胞内Ca2+が中片に比較的高濃度で維持されていますが、SMISが結合すると中片の細胞内Ca2+濃度がさらに上昇します (Watanabe et al., 2011)。細胞内Ca2+濃度の上昇は動物の精子の運動調節で一般に起こり、細胞膜などに存在する輸送タンパク質がこの変化を仲介します。アカハライモリの精子には細胞外Ca2+を細胞内に輸送する、電位依存性Ca2+チャネル(L型とT型)、TRPV4チャネル、NMDA型グルタミン酸受容体、cyclic nucleotide-gated チャネルが存在し、SMISによる精子運動の開始にはT型電位依存性Ca2+チャネルとNMDA型グルタミン酸受容体を介してCa2+が細胞内に流入します(Watanabe &
Takayama-Watanabe, 2014; Takayama-Watanabe et al.,
2015; Endo et al., 2019)。これによって開始される精子運動は、L型電位依存性チャネルを介したCa2+の細胞内への流入により活発化され、ジェリー層を通過するために必要な強度の前進運動が精子に起こります (Takahashi et al., 2013)。このように多種類のチャネルタンパク質が連携した運動調節もSMISによる信号伝達の特徴です。
(6)精子の受精能を維持するメカニズム
細胞膜に多種類のCa2+チャネルをもつことは、動物の精子に共通した特徴ですが、適切なタイミングでチャネルを利用するためにそれらの開口が適切に制御される必要があります。実際、アカハライモリの輸精管に貯蔵された精子を生理的食塩水に曝すと、T型電位依存性Ca2+チャネルか、TRPチャネル、NMDA型グルタミン酸受容体、cyclic nucleotide-gated チャネルが自発的に開口して、先体反応を起こしてしまいます (Kon et al., 2017; 2019)。先体反応はジェリー層の表面で起きることが受精の成立に重要であることから、チャネルの自発的な開口による先体反応は精子の受精能は損ないます。このような先体反応は輸精管で長く貯蔵された精子ほど起こしやすく、また、健常な個体の輸精管では自発的な先体反応はほとんど起こっていませんが、飢餓状態が続いた個体では多くの精子に起こっています。健常個体の輸精管ではCa2+チャネルに働いて自発的な先体反応を抑制するメカニズムが働いており、精子の受精能が保たれているようです。それでは、何が輸精管で精子の受精能を保っているのでしょうか。私たちは現在、この問題の鍵となる分子の探索を進めています。
(7)SMISによる精子の運動調節の起源
アカハライモリのユニークな精子運動開始機構は、どのようにして成立したのでしょうか。このことは、鍵となるSMISが現存する両生類種の多様な生殖様式のそれぞれに関与しているかどうか、また、どのように働くかを比較研究することで明らかにすることができます。私たちは、これまでに有尾目両生類では、淡水中で体外受精をするトウホクサンショウウオ(Ohta et al., 2012) と体内受精をするシリケンイモリ (Sato et al., 2017)、無尾両生類では、樹上受精をするモリアオガエル (Yokoe et al.,
2016)、淡水中で体外受精をするイロワケガエル (Takayama-Watanabe et al., 2012) のジェリー層やそこから派生した泡巣などにSMISが存在することを見出しました。アカハライモリでは、等張の塩類溶液中でSMISによって精子運動が誘導されますが、モリアオガエルではSMISは単独で精子運動が誘導されることは無く、その作用は低塩類溶液の低浸透圧に依存していました。一方、イロワケガエルでは 低浸透圧でもSMIS単独でも同様に精子運動が誘導されました。一方、淡水中で体外受精をするアフリカツメガエルとネッタイツメガエルはSMISをもっていないこともわかりました。SMISは、受精様式を問わず、有尾、および無尾両生類の精子運動調節に広く関わる一方で、SMISに対する精子の応答は受精時に精子が曝される外部環境等に対応して改変されていることを示しています。精子が異なる生殖様式に対して適応的に変化できたことが両生類の生殖様式の多様化と体内受精の獲得の主要な要因であったと考えられます。それでは、SMISは進化的に両生類で獲得されたのでしょうか、それとも魚類で既に獲得されていたのでしょうか。これまでのところ、SMISは両生類以外の動物では発見さてれていませんが、この問題は今後の検討課題となっています。
参照文献
#
Duellman WE, Trueb L. 1994. Biology of amphibians. Baltimore: Johns Hopkins
University Press.
# Inoda T, Morisawa, M. 1987.
Effect of osmolality on the initiation of sperm motility in Xenopus laevis.
Comp. Biochem. Physiol. 88A: 539–542.
#
Sasaki T, Kamimura S, Takai H, Watanabe A, Onitake K.
2002. The activity for the induction of sperm acrosome reaction localises in the outer layers and exists in the high
molecular weight components of egg-jelly of the newt, Cynops
pyrrhogaster. Zygote 10: 1-9.
# Itoh
T, Kamimura S, Watanabe A, Onitake K. 2002. Egg-jelly
structure promotes efficiency of internal fertilization in the newt, Cynops pyrrhogaster. J. Exp.
Zool. 292: 314-322.
# Nakai
S, Watanabe A, Onitake K. 1999. Sperm surface
heparin/heparan sulfate is responsible for sperm binding to the uterine
envelope in the newt, Cynops pyrrhogaster.
Develop. Growth Differ. 41: 101-107.
#
Campanella, C, Carotenuto R, Infante V, Maturi G, Atripalde
U. 1997. Sperm-egg interaction in the painted frog (Discoglossus
pictus): an ultrastructural study. Mol. Reprod. Dev.
47: 323-333.
#
Yoshizaki N, Katagiri C. 1982. Acrosome reaction in sperm of the
toad, Bufo bufo japonicus. Gamete Res. 6:
343-352.
#
Takahashi S, Nakazawa H, Watanabe A, Onitake K. 2006.
The outermost layer of egg-jelly is crucial to successful fertilization in the
newt, Cynops pyrrhogaster.
J. Exp. Zool. 305A: 1010-1017.
#
Watanabe A, Fukutomi K, Kubo H, Ohta M, Takayama-Watanabe E, Onitake K. 2009. Identification of egg-jelly substances
triggering sperm acrosome reaction in the newt, Cynops
pyrrhogaster. Mol. Reprod.
Dev.79: 399-406.
#
Watanabe T, Kubo H, Takeshima S, Nakagawa M, Ohta M, Kamimura S,
Takayama-Watanabe E, Watanabe A, Onitake K. 2010.
Identification of the sperm motility-initiating substance in the newt, Cynops pyrrhogaster, and its possible relation with the acrosome reaction during
internal fertilization. Int. J. Dev.
Biol. 54: 591-597.
# Yokoe M, Sano M, Shibata H, Shibata D,
Takayama-Watanabe E, Inaba K, Watanabe A. 2014. Sperm proteases that may be involved in the initiation of sperm
motility in the newt, Cynops pyrrhogaster.
Int. J. Mol. Sci. 15: 15210-15224.
# Yokoe M, Takayama-Watanabe E, Saito Y, Kutsuzawa
M, Fujita K, Ochi H, Nakauchi Y, Watanabe A. 2016. A novel cysteine knot
protein for enhancing sperm motility that might facilitate the evolution of
internal fertilization in amphibians. PLOS ONE 11: e0160445.
#
Mizuno J, Watanabe A, Onitake K. 1999. Initiation of
sperm motility in the newt, Cynops pyrrhogaster, is induced by a heat-stable component of
egg-jelly. Zygote 7: 329-334.
#
Watanabe A, Takayama-WatanabeE, Vines CA, Cherr GN. 2011. Sperm motility-initiating substance in newt
egg-jelly induces differential initiation of sperm motility based on sperm
intracellular calcium levels. Develop. Growth Differ. 53: 9-17.
#
Watanabe A, Takayama-Watanabe E. 2014. In silico identification of the genes
for sperm-egg interaction in the internal fertilization of the newt Cynops pyrrhogaster. Int. J. Dev.
Biol. 58: 873-879.
# Takahashi T, Kutsuzawa M, Shiba K, Takayama-Watanabe E, Inaba K,
Watanabe A. 2013. Distinct Ca2+ channels maintain a high motility
state of the sperm that may be needed for penetration of egg jelly of the newt,
Cynops pyrrhogaster.
Develop. Growth Differ. 55: 657-667.
#
Takayama-Watanabe E, Ochiai H, Tanino S, Watanabe A. 2015. Contribution of
different Ca2+ channels to the acrosome reaction-mediated initiation
of sperm motility in the newt Cynops pyrrhogaster. Zygote 23:
342-351.
# Endo D, Kon S, Sato T, Toyama F, Katsura
Y, Nakauchi Y, Takayama-Watanabe E, Watanabe A. 2019.
NMDA-type glutamate receptors mediate the acrosome reaction and
motility initiation in newt sperm. Mol. Reprod. Dev.
86: 1106-1115. doi: 10.1002/mrd.23225.
# Kon
S, Sato T, Endo D, Takahashi T, Takaku A, Nakauchi Y,
Toyama F, Meyer-Rochow VB, Takayama-Watanabe E, Watanabe A. 2017. Sperm storage
influences the potential for spontaneous acrosome reaction of the sperm in the
newt Cynops pyrrhogaster.
Mol. Reprod. Dev. 84: 1314-1322. DOI:
10.1002/mrd.22932.
# Kon
S, Takaku A, Toyama F, Takayama-Watanabe E, Watanabe
A. 2019. Acrosome reaction inducing substance triggers two different pathways
of sperm intracellular signaling in newt fertilization. Int. J. Dev. Biol. in
press.
# Ohta
M, Kubo H, Nakauchi Y, Watanabe A. 2010. Sperm motility-initiating activity in
the egg jelly of the externally-fertilizing urodele
amphibian, Hynobius lichenatus.
Zool. Sci. 27: 875-879.
# Takayama-Watanabe E,
Campanella C, Kubo H, Watanabe A. 2012. Sperm motility initiation by egg jelly
of the anuran, Discoglossus pictus may be mediated by
sperm motility-initiating substance of the internally-fertilizing
newt, Cynops pyrrhogaster.
Zygote 20: 417-422.